One of the best documented examples of feedback is that which governs the center-surround receptive field organization of retinal neurons. There is abundant evidence that the earliest stage at which feedback occurs is at the first synapse in the visual pathway, i.e., between the axon terminals of photoreceptors and the dendritic processes of horizontal cells both for cones and for rods. Although extensively investigated in many vertebrate species, there is still no general agreement as to the mechanism that mediates feedback. Nevertheless, two appealing, but very different, views on how feedback modulates the Ca2+current in cones have emerged from recent studies on lower vertebrates. According to the Lomitapide Mesylate hemichannel hypothesis, surround illumination causes 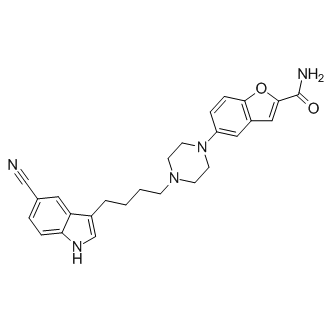 the horizontal cell to hyperpolarize, thus leading to an increase in the current flowing through both the hemichannels and the glutamategated channels. This current flow produces a voltage drop along the high resistance path of the synaptic cleft, thereby shifting the Ca2+ current in the cones to more negative potentials and enhances glutamate release from the cone terminal. An alternative view of the mechanism that mediates feedback in the distal retina was put forth by Hirasawa and Kaneko. They found that the feedback-induced shift of the Ca2+ current in cones was inhibited when extracellular proton fluctuations were stabilized by a high concentration of the pH buffer HEPES; the high concentration of HEPES also reduced light-induced surround effects in bipolar cells. These data together with evidence that increasing extracellular pH shifts the L-type Ca2+ current toward negative potentials led them to conclude that protons regulate the horizontal cell-to-cone feedback pathway. In the present study, we performed a series of electrophysiological experiments designed to test the pH-mediated feedback mechanism and to determine whether the effects of extracellular pH buffering can also be evaluated in terms of the ephaptic feedback hypothesis. A major feature of these experiments is the use of a series of pharmacological agents that change the intracellular pH or the extracellular pH buffering. Our 3,4,5-Trimethoxyphenylacetic acid experimental findings suggest that artificial pH buffers, on which several key arguments supporting the pH hypothesis are based, induce intracellular acidification in addition to clamping the extracellular pH. Because hemichannels are inhibited by intracellular acidification, these experiments do not seem to discriminate between a pH-mediated or a hemichannel-mediated mechanism. Independent experiments to further test the pH hypothesis failed to generate support for the pH-mediated mechanism. Moreover, the experimental results concerning the endogenous pH buffering system, and the computational analysis, are in line with an ephaptic feedback pathway that operates through both hemichannels and glutamate-gated channels. The fact that increasing the pH buffering capacity of the extracellular milieu with HEPES suppressed the light-induced feedback signal raises the question of whether the feedback or the feedforward signal was affected. Any decrease in the feedforward signal will obviously lead to a decrease in the light-induced feedback-signal, because the feedback pathway will receive less input. Because our findings indicate that HEPES reduced the light response of horizontal cells.
the horizontal cell to hyperpolarize, thus leading to an increase in the current flowing through both the hemichannels and the glutamategated channels. This current flow produces a voltage drop along the high resistance path of the synaptic cleft, thereby shifting the Ca2+ current in the cones to more negative potentials and enhances glutamate release from the cone terminal. An alternative view of the mechanism that mediates feedback in the distal retina was put forth by Hirasawa and Kaneko. They found that the feedback-induced shift of the Ca2+ current in cones was inhibited when extracellular proton fluctuations were stabilized by a high concentration of the pH buffer HEPES; the high concentration of HEPES also reduced light-induced surround effects in bipolar cells. These data together with evidence that increasing extracellular pH shifts the L-type Ca2+ current toward negative potentials led them to conclude that protons regulate the horizontal cell-to-cone feedback pathway. In the present study, we performed a series of electrophysiological experiments designed to test the pH-mediated feedback mechanism and to determine whether the effects of extracellular pH buffering can also be evaluated in terms of the ephaptic feedback hypothesis. A major feature of these experiments is the use of a series of pharmacological agents that change the intracellular pH or the extracellular pH buffering. Our 3,4,5-Trimethoxyphenylacetic acid experimental findings suggest that artificial pH buffers, on which several key arguments supporting the pH hypothesis are based, induce intracellular acidification in addition to clamping the extracellular pH. Because hemichannels are inhibited by intracellular acidification, these experiments do not seem to discriminate between a pH-mediated or a hemichannel-mediated mechanism. Independent experiments to further test the pH hypothesis failed to generate support for the pH-mediated mechanism. Moreover, the experimental results concerning the endogenous pH buffering system, and the computational analysis, are in line with an ephaptic feedback pathway that operates through both hemichannels and glutamate-gated channels. The fact that increasing the pH buffering capacity of the extracellular milieu with HEPES suppressed the light-induced feedback signal raises the question of whether the feedback or the feedforward signal was affected. Any decrease in the feedforward signal will obviously lead to a decrease in the light-induced feedback-signal, because the feedback pathway will receive less input. Because our findings indicate that HEPES reduced the light response of horizontal cells.
Month: June 2019
Normal expression inhibits the translation of several cytok especially prominent
Represents a Lomitapide Mesylate strong point of shared biology between human ASD and the MIA model. One of the first structural brain abnormalities found in children with ASD was hypoplasia of the cerebellar vermis that preferentially affected lobules VI and VII. This has also been documented in adults with Fragile X Orbifloxacin Syndrome. Cerebellar Purkinje cells are large, fast-spiking, GABAergic, inhibitory neurons that are particularly sensitive to bioenergetic supply and demand problems, and to toxic exposures. Our finding of preserved cerebellar Purkinje cell numbers at 16 weeks of age in the MIA model with antipurinergic therapy supports the notion that the rate of postnatal Purkinje cell loss is dynamic and can be regulated by environmental factors. In the MIA mouse model, antipurinergic therapy slows the rate of Purkinje cell loss from 6 to 16 weeks of age. Like human autism spectrum disorders, the MIA mouse model of ASD has both core behavioral abnormalities, and multisystem comorbidities that emerge as a consequence of underlying metabolic disturbances. Our results support the paradigm that all of the observed metabolic disturbances in this model are a manifestation of the conserved cell danger response. The CDR therefore lies at or near the root cause of the neurodevelopmental and biochemical abnormalities that characterize the ASD-like features in this model. Extracellular ATP is a mitokine and well-known danger signal that we hypothesized initiates and sustains the cellular danger response in autism spectrum disorders. In related studies we found that direct systemic injection of nucleotides like ATP and ADP caused rapid hypothermia 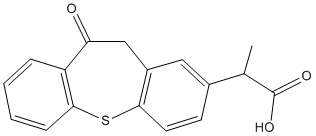 by decreasing mitochondrial oxygen consumption and tissue oxygen demand. Hypothermia from systemic nucleotide injection has been studied in the fields of torpor and hibernation physiology. We found that a convenient marker of the persistent cellular danger response in the poly model is relative hypothermia of about 0.5uC. Hypothermia was associated with an increase in the maximal enzymatic rates, but not the mass, of brain mitochondrial respiratory chain complexes I and IV. Treatment with suramin decreased brain mitochondrial activity to normal, increased the whole body oxygen consumption in the MIA animals, and increased the body temperature to normal. The combination of higher mitochondrial electron transport activities measured in vitro and decreased basal oxygen consumption measured in vivo implies a novel increase in mitochondrial coupling efficiency and increased reserve capacity in ASD that is similar to that seen with exercise training. We did not further investigate this phenomenon in this study. Purinergic P2Y2 receptors and their phosphorylated effectors, ERK1/2 and CAMKII, are downregulated by chronic nucleotide stimulation in a process that leads to desensitization. Our finding of downregulation of these purinergic receptors and their effectors is strong evidence for chronically elevated purinergic signaling in the poly model. Together, these findings are consistent with the notion that hyperpurinergia is a causal factor that initiates and maintains the cellular danger response in the MIA model of ASD. Suramin treatment corrected both the hyperpurinergia and the multisystem abnormalities in this model. We quantified the expression of the Fragile X protein in cerebral synaptosomes because deficiency is a cause of autism spectrum disorders.
by decreasing mitochondrial oxygen consumption and tissue oxygen demand. Hypothermia from systemic nucleotide injection has been studied in the fields of torpor and hibernation physiology. We found that a convenient marker of the persistent cellular danger response in the poly model is relative hypothermia of about 0.5uC. Hypothermia was associated with an increase in the maximal enzymatic rates, but not the mass, of brain mitochondrial respiratory chain complexes I and IV. Treatment with suramin decreased brain mitochondrial activity to normal, increased the whole body oxygen consumption in the MIA animals, and increased the body temperature to normal. The combination of higher mitochondrial electron transport activities measured in vitro and decreased basal oxygen consumption measured in vivo implies a novel increase in mitochondrial coupling efficiency and increased reserve capacity in ASD that is similar to that seen with exercise training. We did not further investigate this phenomenon in this study. Purinergic P2Y2 receptors and their phosphorylated effectors, ERK1/2 and CAMKII, are downregulated by chronic nucleotide stimulation in a process that leads to desensitization. Our finding of downregulation of these purinergic receptors and their effectors is strong evidence for chronically elevated purinergic signaling in the poly model. Together, these findings are consistent with the notion that hyperpurinergia is a causal factor that initiates and maintains the cellular danger response in the MIA model of ASD. Suramin treatment corrected both the hyperpurinergia and the multisystem abnormalities in this model. We quantified the expression of the Fragile X protein in cerebral synaptosomes because deficiency is a cause of autism spectrum disorders.
Which is primarily due to the presence of multiple alleles of various number of tandem repeats
However, the presence of the VNTR, as well as the cDNA sequence within the PTS domain, is not highly conserved evolutionarily, highlighting the distinct possibility that broad functional differences exist between species. Our data indicate that the PTS domain of the LOUREIRIN-B chicken MUC2 protein contains a vastly different repeat structure than the human protein. Although the chicken PTS region is shorter, the central repeat motif is 69 amino acids in length and shows very little identity with the human motif. Recent in vitro studies using human intestinal cells demonstrated that the intestinal mucins isolated from chicken were detrimental to the proliferation of Campylobacter jejuni, an infectious bacteria causing acute gastroenteritis in humans but not in chicken. In addition these studies demonstrated that the chicken mucins attenuated the invasiveness of Campylobacter jejuni, suggesting that differences in mucin protein sequence or structure between humans and chicken could account for the differences in susceptibility to infection. Alternatively, the functional differences between human and chicken may imply species-specific divergence in intestinal mucus composition and/or structures. This could also occur through differences in posttranslational modifications of the human and chicken proteins. Outside of the PTS domain, the human and chicken MUC2 proteins share large blocks of highly conserved sequences, strongly suggesting that this variable PTS region could account for the phenotypic differences. Plausibly, MUC2 is of 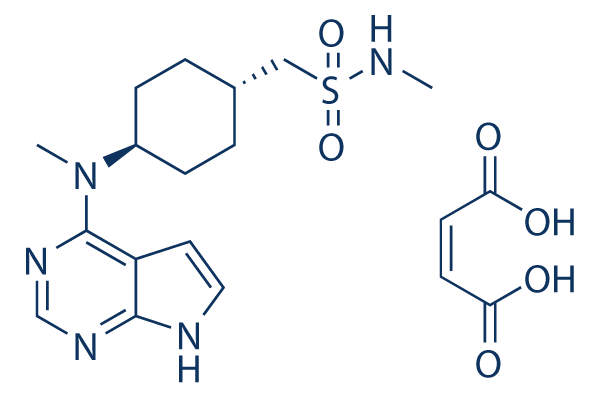 utmost importance, as the functionality of intestinal mucus was proposed to rely primarily on MUC2 encoded mucins. Therefore, the full understanding of the functional divergence and prognostic implications of chicken mucins compared to their mammalian orthologues necessitates identification and comparisons of the gene sequences across species. Although identification of new MUC family members is ongoing, sequencing of most MUCIN genes is hampered due to the highly complex PTS cassettes clustered throughout the gene, and several gaps still remain in mouse and human family members. In the case of the secretory mucins, this can largely be accounted for by the large, frequently repetitive PTS region. The presence of several different polymorphic elements in many of the MUCIN genes hinders annotation efforts at the gene and protein levels, and could even hamper the understanding of the biological Orbifloxacin significance and disease associations of the diverse family members. In the endoplasmic reticulum, MUC2 forms disulfide-linked dimers via the VWD domains of the amino-terminus, while the CT knot in the carboxy-terminus supports disulfide-linked trimerization in the trans-Golgi network. CysD domains exert non-covalent cross-linkages in the MUC2 gel formation process, likely contributing to tertiary structure and determination of the pore size of the mucus network. Chicken may plausibly carry more CysD domains than that of human, which may suggest that the polymeric net-like structure contains smaller pores in chickens than humans. This could account for differences in innate defense response to pathogens. The conservation of a cationic domain at the C-terminus observed in rodents was not found in chickens. In human MUC2, two different PTS domains have been identified, both of which are located on the same large exon separated by,600 bp. One region consists of repeats that are interrupted in places by 21 to 24 bp segments. The other is composed of an uninterrupted array of a tandem 23-amino acid repeat cassette.
utmost importance, as the functionality of intestinal mucus was proposed to rely primarily on MUC2 encoded mucins. Therefore, the full understanding of the functional divergence and prognostic implications of chicken mucins compared to their mammalian orthologues necessitates identification and comparisons of the gene sequences across species. Although identification of new MUC family members is ongoing, sequencing of most MUCIN genes is hampered due to the highly complex PTS cassettes clustered throughout the gene, and several gaps still remain in mouse and human family members. In the case of the secretory mucins, this can largely be accounted for by the large, frequently repetitive PTS region. The presence of several different polymorphic elements in many of the MUCIN genes hinders annotation efforts at the gene and protein levels, and could even hamper the understanding of the biological Orbifloxacin significance and disease associations of the diverse family members. In the endoplasmic reticulum, MUC2 forms disulfide-linked dimers via the VWD domains of the amino-terminus, while the CT knot in the carboxy-terminus supports disulfide-linked trimerization in the trans-Golgi network. CysD domains exert non-covalent cross-linkages in the MUC2 gel formation process, likely contributing to tertiary structure and determination of the pore size of the mucus network. Chicken may plausibly carry more CysD domains than that of human, which may suggest that the polymeric net-like structure contains smaller pores in chickens than humans. This could account for differences in innate defense response to pathogens. The conservation of a cationic domain at the C-terminus observed in rodents was not found in chickens. In human MUC2, two different PTS domains have been identified, both of which are located on the same large exon separated by,600 bp. One region consists of repeats that are interrupted in places by 21 to 24 bp segments. The other is composed of an uninterrupted array of a tandem 23-amino acid repeat cassette.
Dexamethasone had limited effects on AHR but did attenuate the inflammatory cell influx
Mammalian target of rapamycin signaling occurs downstream of the PI3K-signaling cascade and is known to play a major role in growth/differentiation, cell metabolism, and survival in many different cell types. More recent work has demonstrated an important role for mTOR in T cell proliferation and differentiation. An inhibitor of mTOR, rapamycin, is already used clinically as an immunosuppressant to prevent organ Lomitapide Mesylate rejection after transplantation. In addition, the use of rapamycin in patients suffering from the destructive lung disease, lymphangioleiomyomatosis, has demonstrated promise in its ability to reduce disease symptoms and stabilize lung function. Previously, our lab demonstrated that inhibition of mTOR with rapamycin prevented Mepiroxol allergic asthma in a mouse model induced by exposure to the allergen, house dust mite. In these studies, rapamycin prevented the allergic response and still suppressed many key asthma characteristics after allergic sensitization was established. Although this study showed that mTOR inhibition could suppress allergic asthma early in the disease process, the role of mTOR during allergen reexposure and chronic, established allergic disease remained unclear. The goal of this study was to determine whether inhibition of mTOR with rapamycin would attenuate key characteristics of allergic asthma in two models that addressed chronic/established disease, namely allergen re-exposure and disease progression. In addition to rapamycin, mice were also treated with the steroid, dexamethasone, for comparison purposes since steroids are currently a mainstay therapy for chronic asthma. We hypothesized that rapamycin and dexamethasone would suppress asthma exacerbations during allergen re-exposure and suppress progressive/ongoing allergic disease by inhibiting T cells. To test this hypothesis, mice in protocol one, which was designed to mimic the effects of allergen re-exposure in a previously sensitized individual, were sensitized to HDM by i.p. injection and then exposed to intranasal HDM to induce asthma. Then, after a 6 week rest/recovery period, mice were re-exposed to HDM while being treated with rapamycin or dexamethasone. In protocol two, to address the role of mTOR in chronic/established allergic asthma, mice were exposed to HDM for 6 weeks and treated with rapamycin or dexamethasone from weeks 4 to 6 of the exposure period. Endpoints assessed included allergen specific IgE, AHR, inflammatory cells, goblet cell metaplasia, cytokine/chemokine levels, and T cell numbers. The goal of our study was to determine whether mTOR inhibition 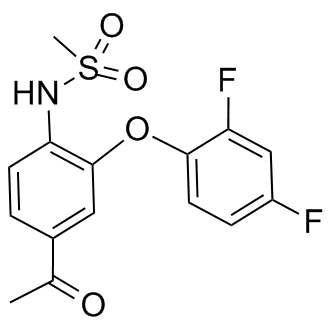 with rapamycin would suppress the key features and mediators of HDM-induced allergic asthma in established asthmatic disease. In addition, we also compared rapamycin to the steroid, dexamethasone, since steroids are currently a mainstay treatment for asthma. In the first protocol, we assessed whether rapamycin or dexamethasone could suppress allergic disease during allergen re-exposure. Although rapamycin suppressed IgE levels, goblet cells, and total lung T cells, it had no effect on AHR or BALF cellularity and IL-4 and eotaxin 1 levels were actually augmented. Dexamethasone suppressed goblet cells and total lung T cells, but had no effect on IgE or AHR and only slightly reduced BALF eosinophilia. Our second protocol assessed whether rapamycin or dexamethasone could reverse or inhibit the progression of asthmatic responses during chronic allergic airway disease. In this model, rapamycin did not suppress AHR or goblet cells and actually augmented inflammatory cell numbers, IL-4 and eotaxin 1 levels in the BALF.
with rapamycin would suppress the key features and mediators of HDM-induced allergic asthma in established asthmatic disease. In addition, we also compared rapamycin to the steroid, dexamethasone, since steroids are currently a mainstay treatment for asthma. In the first protocol, we assessed whether rapamycin or dexamethasone could suppress allergic disease during allergen re-exposure. Although rapamycin suppressed IgE levels, goblet cells, and total lung T cells, it had no effect on AHR or BALF cellularity and IL-4 and eotaxin 1 levels were actually augmented. Dexamethasone suppressed goblet cells and total lung T cells, but had no effect on IgE or AHR and only slightly reduced BALF eosinophilia. Our second protocol assessed whether rapamycin or dexamethasone could reverse or inhibit the progression of asthmatic responses during chronic allergic airway disease. In this model, rapamycin did not suppress AHR or goblet cells and actually augmented inflammatory cell numbers, IL-4 and eotaxin 1 levels in the BALF.
Clearly indicated peripheral nociception for thermal or mechanical sensitivity
The development and/or maintenance of thermal and mechanical hyperalgesia and spontaneous pain in a variety of pain models; EphB1 receptors are necessary for the induction of phosphorylation of the NR2B subunit of the NMDA receptor and are Lomitapide Mesylate involved in the increase of c-fos expression in models of inflammatory pain and tissue injury and in microglial activation following PNL. EphB1 receptors, together with other Eph receptors, play an important role in neuronal development, therefore it was important to establish that EphB1 KO mice were normal in terms of development of nociceptive pathways and nociception, in order to be suitable in models for the study of the modulation of pain processing. This had not been examined in previous studies using these mice. Since pain sensitivity in animal models is measured through behavioural tests dependent on a motor response, it was also important to test na? ��ve KO mice in comparison to WT  littermate in a motor test, particular in light of findings indicating that EphB1 KO exhibit neuronal loss in the substantia nigra. We found no indication that this loss affected the response to acute noxious stimuli using a variety of tests, including thermal and mechanical stimuli, or to the first phase of the formalin test, reflecting acute tissue damage. Gross anatomical abnormalities were also absent from both the DRG and the dorsal horn of the spinal cord. Similarly electrophysiological recording at the cord dorsum, measuring the spatial distribution and amplitudes of the CAPs and FPs following sciatic nerve stimulation, did not reveal any difference between WT and EphB1 KO mice in terms of somatotopic organisation of sensory projections in the spinal cord. Although the presence of developmental defects in glutamatergic synapses in the hippocampus of transgenic EphB2 KO mice remains unclear, it is possible that in single mutants compensatory mechanisms, due to the presence of other EphB receptors, which bind promiscuously to the same ephrinB ligands, may allow a comparatively normal development. We cannot however exclude the presence of subtle changes, for example at the level of the glutamatergic synapses of sensory afferents onto dorsal horn neurons, below the level of detection behaviourally. In this respect our finding of enhanced sural nerve evoked superficial dorsal horn FPs is of relevance in that it indicates such a change in the connectivity of Ab-fibres. Our simplest explanation for this finding draws on the changes Mepiroxol observed early postnatally in the rat where the predominant A-fibre innervation of the superficial dorsal horn at birth undergoes a dramatic, activity-dependent withdrawal of the nerve terminals to deeper layers over the first postnatal weeks. It is proposed here that such functional and structural reorganisation did not occur in EphB1KO mice. Further studies are needed to determine whether inhibitory or excitatory interneurons, projection neurons or both, are involved in this refinement process. Although this change in the connectivity of A fibres in the sural nerve is of considerable interest in view of the guidance role of Eph receptors during development, it had no demonstrable impact on the behavioural tests we employed, or on the gross anatomical connectivity of C and Ad fibres. Another possible explanation for the enhanced synaptic transmission we observed could be a change in the electrophysiological properties of the large diameter Ab-fibre, as described in DRG neurons for a rat model of osteoarthritic pain and more recently in a neuropathic pain model.
littermate in a motor test, particular in light of findings indicating that EphB1 KO exhibit neuronal loss in the substantia nigra. We found no indication that this loss affected the response to acute noxious stimuli using a variety of tests, including thermal and mechanical stimuli, or to the first phase of the formalin test, reflecting acute tissue damage. Gross anatomical abnormalities were also absent from both the DRG and the dorsal horn of the spinal cord. Similarly electrophysiological recording at the cord dorsum, measuring the spatial distribution and amplitudes of the CAPs and FPs following sciatic nerve stimulation, did not reveal any difference between WT and EphB1 KO mice in terms of somatotopic organisation of sensory projections in the spinal cord. Although the presence of developmental defects in glutamatergic synapses in the hippocampus of transgenic EphB2 KO mice remains unclear, it is possible that in single mutants compensatory mechanisms, due to the presence of other EphB receptors, which bind promiscuously to the same ephrinB ligands, may allow a comparatively normal development. We cannot however exclude the presence of subtle changes, for example at the level of the glutamatergic synapses of sensory afferents onto dorsal horn neurons, below the level of detection behaviourally. In this respect our finding of enhanced sural nerve evoked superficial dorsal horn FPs is of relevance in that it indicates such a change in the connectivity of Ab-fibres. Our simplest explanation for this finding draws on the changes Mepiroxol observed early postnatally in the rat where the predominant A-fibre innervation of the superficial dorsal horn at birth undergoes a dramatic, activity-dependent withdrawal of the nerve terminals to deeper layers over the first postnatal weeks. It is proposed here that such functional and structural reorganisation did not occur in EphB1KO mice. Further studies are needed to determine whether inhibitory or excitatory interneurons, projection neurons or both, are involved in this refinement process. Although this change in the connectivity of A fibres in the sural nerve is of considerable interest in view of the guidance role of Eph receptors during development, it had no demonstrable impact on the behavioural tests we employed, or on the gross anatomical connectivity of C and Ad fibres. Another possible explanation for the enhanced synaptic transmission we observed could be a change in the electrophysiological properties of the large diameter Ab-fibre, as described in DRG neurons for a rat model of osteoarthritic pain and more recently in a neuropathic pain model.