An interpretation of these current findings is that in macaque and human cortex there is a larger population of GluA2 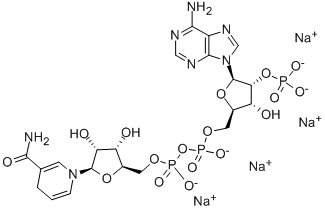 associated with the ER and/or early Golgi cellular compartments. Interestingly, earlier studies have demonstrated that the ER localization of the GluA2 subunit is essential for assembly of AMPA receptor complexes, the exit of assembled receptors from the ER, and forward trafficking to the synaptic membrane. Not detecting N-linked glycans by deglycosylation assays was somewhat surprising, given previous findings that the GluA1 and GluA3 subunits from rat frontal cortex were sensitive to enzymatic glycosylation. Our data suggest either the possibility of a low molecular mass of N-linked glycans on GluA1 and GluA3 subunits that went undetectable by our enzymatic deglycosylation, or alternatively the presence of an unknown modification that confers glycans resistance to enzymatic cleavage. The lectin binding assay is more sensitive due to the enrichment of glycans specific to each lectin, but is not specific to the type of glycosylation. For example, O-linked glycosylation of AMPA receptor subunits is a possibility that could Coptisine-chloride explain the lack of detection of Nglycosylation by Endo H or PNGase F, and potentially consistent with the presence of glycosylated GluA1 and GluA3 subunits that we found by lectin binding. The modest differences that we found in glycosylation of AMPA receptor subunits in the human may Loganin reflect intrinsic differences in biosynthesis, processing, trafficking, or interaction of the receptor subunits with cellular and extracellular partners. Recent evolutionary findings suggest that within each phylogenetic branch, the extracellular glycoproteome coevolved by adapting to extracellular environmental cues specific to development, growth, and organ formation. Extracellular N-glycosylated proteins appear to have evolved at much faster rate than intracellular glycoproteins, except for extracellular N-glycosylated Asn. Interestingly, the nonN-glycosylated Asn evolved at significantly higher rate than Nglycosylated, suggesting a significant evolutionary pressure to retain N-glycosylated Asn residues. Both the intracellular and extracellular glycoproteomes are essential for cell survival, while balanced excitatory synaptic transmission is necessary for higher functioning. Genetic information for glycosylation is conserved, yet the versatility of post-translational modifications varies among species, notably in the terminal glycans. This suggests that AMPA receptors exit the Golgi after terminal glycosylation driven by species-specific cellular and environmental pressures. In summary, this study compares the glycosylation pa ern of AMPA receptor subunits in the frontal cortex across different mammalian species. We found that all four AMPA receptor subunits are glycosylated, but also demonstrated that there are differences in glycosylation between different subunits as well as modest differences in glycosylation of homologous subunits between different species. Nasopharyngeal carcinoma is a common malignant disease in Southeast Asia, with an annual incidence of 30�C80 per 10,000. Approximately 80% of patients present with advanced disease at first diagnosis as a result of its silent, deep-seated location and non-specific symptoms.
associated with the ER and/or early Golgi cellular compartments. Interestingly, earlier studies have demonstrated that the ER localization of the GluA2 subunit is essential for assembly of AMPA receptor complexes, the exit of assembled receptors from the ER, and forward trafficking to the synaptic membrane. Not detecting N-linked glycans by deglycosylation assays was somewhat surprising, given previous findings that the GluA1 and GluA3 subunits from rat frontal cortex were sensitive to enzymatic glycosylation. Our data suggest either the possibility of a low molecular mass of N-linked glycans on GluA1 and GluA3 subunits that went undetectable by our enzymatic deglycosylation, or alternatively the presence of an unknown modification that confers glycans resistance to enzymatic cleavage. The lectin binding assay is more sensitive due to the enrichment of glycans specific to each lectin, but is not specific to the type of glycosylation. For example, O-linked glycosylation of AMPA receptor subunits is a possibility that could Coptisine-chloride explain the lack of detection of Nglycosylation by Endo H or PNGase F, and potentially consistent with the presence of glycosylated GluA1 and GluA3 subunits that we found by lectin binding. The modest differences that we found in glycosylation of AMPA receptor subunits in the human may Loganin reflect intrinsic differences in biosynthesis, processing, trafficking, or interaction of the receptor subunits with cellular and extracellular partners. Recent evolutionary findings suggest that within each phylogenetic branch, the extracellular glycoproteome coevolved by adapting to extracellular environmental cues specific to development, growth, and organ formation. Extracellular N-glycosylated proteins appear to have evolved at much faster rate than intracellular glycoproteins, except for extracellular N-glycosylated Asn. Interestingly, the nonN-glycosylated Asn evolved at significantly higher rate than Nglycosylated, suggesting a significant evolutionary pressure to retain N-glycosylated Asn residues. Both the intracellular and extracellular glycoproteomes are essential for cell survival, while balanced excitatory synaptic transmission is necessary for higher functioning. Genetic information for glycosylation is conserved, yet the versatility of post-translational modifications varies among species, notably in the terminal glycans. This suggests that AMPA receptors exit the Golgi after terminal glycosylation driven by species-specific cellular and environmental pressures. In summary, this study compares the glycosylation pa ern of AMPA receptor subunits in the frontal cortex across different mammalian species. We found that all four AMPA receptor subunits are glycosylated, but also demonstrated that there are differences in glycosylation between different subunits as well as modest differences in glycosylation of homologous subunits between different species. Nasopharyngeal carcinoma is a common malignant disease in Southeast Asia, with an annual incidence of 30�C80 per 10,000. Approximately 80% of patients present with advanced disease at first diagnosis as a result of its silent, deep-seated location and non-specific symptoms.